
_-_Fig._241.gif)




.JPG)


Discovery
During the 1970s, two fossil hunting teams began uncovering evidence of ancient human ancestors in east Africa. In the north, the International Afar Research Expedition (IARE) led by the geologist Maurice Taieb and the anthropologists Yves Coppens and Donald Johanson began collecting fossil remains at Hadar in the Afar region of Ethiopia in 1972. Meanwhile, 1,500 km to the south another team led by Mary Leakey began exploring the area of Laetoli in Tanzania in 1974. Both expeditions proved to be highly fruitful.
In 1973 the Afar expedition found a perfectly articulated knee joint (AL 129 1a/b), which seemed to indicate that hominins had been walking on their hind legs from at least 3½ million years ago. In the following year they discovered the well-preserved skeleton known to the world as ‘Lucy’ and in the year after that they recovered 250 skeletal elements, the very scrambled remains of at least 17individuals, representing at least 9 adults, 3 adolescents, and 5  juveniles, collectively known as the ‘First Family’. Although less than half of Lucy’s skeleton (A.L. 288-1) was recovered, it is possible to reconstruct nearly 80% of it by using lateral symmetry. The First Family included specimens from both sexes and all ages to provide a wide range of comparative material to allow scholars to build up a broader picture of the species as a whole. Meanwhile, at about the same time in Laetoli, Leakey’s team recovered a mandible (LH 4) belonging to the same species. In 1976 they discovered unique sets of footprints preserved in volcanic ash that were clearly made by a hominin, and one that was contemporary with the fossil remains.
juveniles, collectively known as the ‘First Family’. Although less than half of Lucy’s skeleton (A.L. 288-1) was recovered, it is possible to reconstruct nearly 80% of it by using lateral symmetry. The First Family included specimens from both sexes and all ages to provide a wide range of comparative material to allow scholars to build up a broader picture of the species as a whole. Meanwhile, at about the same time in Laetoli, Leakey’s team recovered a mandible (LH 4) belonging to the same species. In 1976 they discovered unique sets of footprints preserved in volcanic ash that were clearly made by a hominin, and one that was contemporary with the fossil remains.
The remains discovered at the two sites were very similar to each other and of a similar age but were unlike any species known at that time. A new species name, Australopithecus afarensis, was therefore created for them in 1978 and, because dentition is the most useful way of distinguishing early hominin species, LH 4 was designated as the ‘type’ specimen. The species was named for the Afar region where the first discoveries were made. It has not been found outside of East Africa so far. A. afarensis is considered by many to by the common ancestor of all later australopithecines (including Paranthropus) and therefore all humans. However, this assumption is not universally held and Praeanthropus has been suggested as an alternative genus.
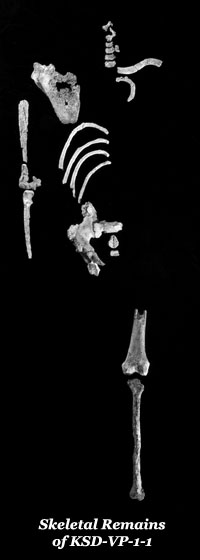
Since the 1970s, a large number of fossils have been discovered up and down the Rift Valley—hundreds of bones representing dozens of individuals and most parts of the skeleton. Most notably they include the skull and partial skeleton of a juvenile (DIK-1-1) at Dikika in 2000 by a team led by Zeresenay Alemseged, and the partial skeleton (KSD-VP-1/1) of a male nicknamed Kadanuumuu (‘big man’ in the local language) consisting of postcranial bones found in Afar in 2005 by a team led by Yohannes Haile-Selassie.
In 2009 a partial foot (BRT-VP-2/73) was found in the Burtele region of Ethiopia. Uniquely for a hominin, it had an apelike prehensile big toe. A case for it belonging to a different australopithecine species was opened in 2011 with the discovery nearby of jaw fragments by Dr. Haile-Selassie. The dentition is different so it has been given its own classification, Australopithecus deyiremeda. The discovery of mandible fragments and a tooth some 2,500 kilometres to the west of the Rift Valley in central Chad. These were identified as yet another contested species, Au. bahrelghazali. Yet another new species (a new genus, in fact) was identified at Lomekwi by Lake Turkana in northern Kenya.
The location of the finds is a clear indication of how widespread and varied australopithecines had become in such a relatively short time. The variety in size, shape and configuration of the evidence has caused a bit of a stir in the scholarly world and there has been considerable discussion as to how many species are involved—a single one, with variations, or a number of separate ones.

Hadar deposits looking South. Afar, Ethiopia
Dating
The individual fossils have each been dated by a wide variety of methods— potassium-argon, fission-track, palaeomagnetism, biostratigraphy— and all of them fall into the period from about 3.7-3 million years ago. This places them in about the middle part of the Pliocene Epoch.
morphology
Although less than half of Lucy’s skeleton (A.L. 288-1) was recovered, it is possible to reconstruct nearly 80% of it by using lateral symmetry. She was estimated to be about 110 cm tall but Kadanuumuu, was a good half metre taller. The weight differences are in the same range, about 3:2—males weighing in at about 45 kg and females just shy of 30 kg, if you assume they had human rather than ape-like proportions. The inclination has been to attribute this to sexual dimorphism or to the age of the particular specimen but now it is apparent that, bearing in mind that the samples range over the better part of a million years, the main factor is time with the earlier specimens being smaller and the later ones larger.
The Skull
None of the fossils collected in the 1970s include anything like a complete skull, so for a long time any study of the skull was based on composite reconstructions. However, a number of fairly complete examples, which include the cranium and the associated mandible, have turned up since then.The largest is AL 444-2, which belonged to a rather elderly male if his badly worn teeth are any indication, had a brain capacity of something like 550 cc. Another belonged to a younger, smaller female (AL 822-1) whose brain was evidently smaller (although she did have a much better set of teeth).
The braincase was smaller than that of Au. africanus, ranging from an estimated 365-525 cc or thereabouts for adults—an average of about 445 cc. Infant brains (two and three year olds) in the 275-315 cc range would seem to indicate that brain growth took longer than with apes but not quite as slow as among modern humans. Although there is no direct association between skulls with recorded endocranial volumes and postcranial remains, it may well be that larger brains simply reflect larger bodies (as is the case with modern humans). It has also been suggested that modification and enlargement of some parts of the cerebral cortex, the outermost part of the brain and key to processing information, were involved but this is far from certain.
The vault of the skull was domed, as it is among later hominins, and the cranial floor (the base of the skull) was relatively flat. But, as is the case with all australopithecines, there were still many ape-like features—a low, sloping forehead, prominent brow ridges and a prognathic lower face. AL 444-2 had nuchal crests along the sides of the skull and a sagittal crest  running lengthwise along the top—as do many apes, particularly the males. They were for the attachment of the powerful temporalis muscles that worked their jaws. In the case of the Hadar specimens, the sagittal crest, which is located only on the back of the skull, is not particularly large and is about the same in both males and females. These ‘ectocranial’ features, along with the presence of hollow spaces within the bone (pneumatisation), mean that the skull is relatively large for the size of the brain case. As far as the mandible is concerned, there is a considerable range of size but the shape is more or less the same, with some variation between the sexes. The chin is larger and firmer than that of apes, which tends to be recessive, sharing similarities both with later humans as well as robust australopithecines.
running lengthwise along the top—as do many apes, particularly the males. They were for the attachment of the powerful temporalis muscles that worked their jaws. In the case of the Hadar specimens, the sagittal crest, which is located only on the back of the skull, is not particularly large and is about the same in both males and females. These ‘ectocranial’ features, along with the presence of hollow spaces within the bone (pneumatisation), mean that the skull is relatively large for the size of the brain case. As far as the mandible is concerned, there is a considerable range of size but the shape is more or less the same, with some variation between the sexes. The chin is larger and firmer than that of apes, which tends to be recessive, sharing similarities both with later humans as well as robust australopithecines.
Elsewhere in the skull, there are slight changes to the palate (which is somewhat shallower than is normally the case among australopithecines), to the mandibular fossa (where the lower jaw is hinged), and to the mastoid process which is located behind the ear, where the neck muscles are connected. In general, these features were still much more apelike than human but that does not apply to the foramen magnum (the opening through which the spinal chord connects to the brain), which is centrally located and faces downwards. This feature is essential for bipedal locomotion because it keeps the skull on top of the spine in the vertical position.
Dentition: Because jaws and teeth are far more likely to survive than any other part of the skeleton, they are the principle means of distinguishing one taxon from another. In general, the dentition of Au. afarensis falls between that of its predecessors (A. anamensis, etc.) and its successors (A. africanus and Homo), and in most respects is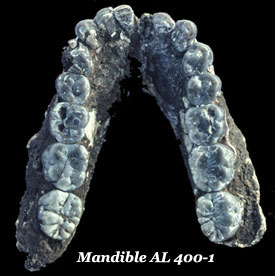 more apelike than human. Unlike humans whose dental arcade has a parabolic arrangement, the Lucy’s teeth have a long, narrow and straight-sided pattern similar to that of apes, except that (in some cases) the hindmost molar is set a little inwards which gives the array a slight curve.
more apelike than human. Unlike humans whose dental arcade has a parabolic arrangement, the Lucy’s teeth have a long, narrow and straight-sided pattern similar to that of apes, except that (in some cases) the hindmost molar is set a little inwards which gives the array a slight curve.
The canines are conical rather than flat, which is a primitive feature, but are smaller—about the same size as Au. anamensis—but not as small as later hominins. There is little difference in size between males and females which suggests that competition among the former for reproductive rights was less intense and the arrangements were less exclusive. The spaces (diastema) needed to accommodate them in the upper and lower jaw are still present but are much smaller and were not continually being honed by rubbing against the lower premolar.
Continuing a trend among australopithecines towards ‘megadonty’, the premolars and the molars are noticeably larger than among the apes, with higher crowns and thicker enamel—capable of plenty of hard chewing and grinding. Like humans, the upper surface of the molars was arranged in a Y-shaped pattern of grooves and raised points known as cusps. The premolars had begun to serve much the same function as the molars but only had two cusps (bicuspid) and there was an apparent trend over time towards a single root rather than two.
Post-Cranial
Our main evidence for the reconstruction of the post-cranial skeleton of the species comes from three discoveries—the remains of AL 288-1 (‘Lucy’), KSD-VP-1/1 (‘Kadanuumuu’) and DIK-1-1 (‘Selam’ or ‘Lucy’s Baby’). Also important (although its classification is disputed) is BRT-VP-2/73 (the ‘Burtele foot’). The remains are far from complete and, in the case of DIK-1-1, have not been completely extricated from their matrix.
In general, like other australopiths, Au. afarensis shares some features with modern humans and others with the rest of the great apes. Unlike the skull, however, most of them are clearly moving toward the human-like. Compared to more modern humans, the arms are longer than the legs, but the difference is not as noticeable as it is with most of the other apes. The lower body was designed to support the weight of a creature used to walking on two feet, while the shoulders and arms are similar to those of orangutans, who spend much of their time moving about in the trees. How much of this is due to inherited traits from their last common ancestor is open to question.
Torso
Ribcage: A number of ribs were included in the skeletal remains of KSD-VP-1/1 and AL 288-1 which show that Au. afarensis had a bell-shaped ribcage much like those of other apes, rather than the barrel-shaped one of later humans—although it was trending in that direction over the lifetime of the species.
Spine: The construction of the spine was also becoming more human, although the evidence is rather sketchy. The vertebral column of an ape runs in a more or less straight line from the head to the coccyx at an approximately 45 degree angle. By contrast the human spine, so the extra weight it has to carry is evenly distributed above the legs, is S-shaped. The cervical and lumbar regions bend backwards (lordosis) while the thoracic and sacral regions curve forwards. No complete spine has so far been recovered but at least one specimen (AL 333-73) shows evidence of ‘wedging’ in the body of the lumbar vertebrae. Wedge-shaped vertebrae indicate a high likelihood of lumbar lordosis. The lack of complete examples also makes it is impossible to say for certain, whether Au. afarensis had four lumbar vertebrae (like apes) or five (like humans).

Skeletal Remains of DIK-1-1 (‘Selam’) © Zeray Alemseged
The cervical (neck) vertebrae got thicker, which probably had to do with their posture and stability of the head. There is evidence in some specimens of a slight bulging of the vertebrae at the base of the neck, the area of the brachial plexus in humans, the seat of the network of nerves that control most of the muscles of the arms and hands. It is possible that this has something to do with advanced motor functions such as manual dexterity but this far from proven and is not observable in all specimens. There was the typical human complement of 7 cervical and 12 thoracic vertebrae (the ones where the ribs attach) in DIK-1-1, rather than 13 as with apes. Unfortunately, only two lumbar vertebrae were recovered and the sacrum was entirely missing.
Shoulders and Arms
Pectoral Girdle: The best preserved pectoral girdle (‘shoulders’) belongs to the juvenile DIK-1-1 and consists of both clavicles as well as the left scapula. A fragment of Lucy’s right scapula (the glenoid and a bit of the lateral or axillary border) also survives. The glenoid fossa (socket) of the scapula still faces slightly 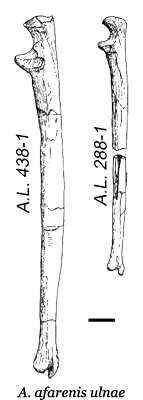 upwards, like an ape’s, rather than to the side like ours. This obviously helps in climbing and hanging from overhead branches. The clavicle (collarbone) retains the S-shape characteristic of apes and would have given the same sort of ‘shrugged shoulder’ appearance.
upwards, like an ape’s, rather than to the side like ours. This obviously helps in climbing and hanging from overhead branches. The clavicle (collarbone) retains the S-shape characteristic of apes and would have given the same sort of ‘shrugged shoulder’ appearance.
Humerus: The relative length of the upper arm compared to the thigh bone (the humerofemoral index) indicates that their arms were proportionally longer than their legs compared to us, but the difference was less pronounced than among apes and earlier hominins, such as Orrorin and Ardipithecus. The head of the humerus is elliptical rather than spheroid as with the great apes and is relatively shorter. From the marks of the muscle attachments it is clear that Au. afarensis had very strong arms.
Radius & Ulna: There is little to be said about the forearm. There are apelike and humanlike features of the radius and ulna and the wrist seems less flexible. The brachial index (the ratio of the length of the forearm to the upper  arm) is also inconclusive, falling within the range of the African apes and later humans. As the illustration (left) shows, there was considerable variation in the length of the arm.
arm) is also inconclusive, falling within the range of the African apes and later humans. As the illustration (left) shows, there was considerable variation in the length of the arm.
The Hand: Reconstruction of the hand is based on an assortment of disconnected bones belonging to the AL 333 collection (the ‘First Family’). It represents a mixture of primitive and derived features (features not found in the common ancestor of all hominins). The primitive ones include curved phalanges and prominent ligament and muscle attachments, which would have given them stronger flexion of the fingers, very useful for brachiation. However, it must be noted that the relative lengths of the AL 438-1 metacarpals is similar to that of humans—that of the thumb was longer than the rest. Possibly this allowed for the sort of precision grip that enabled them to make and use stone tools.
Pelvis & Lower Limbs
Pelvis: As with most humans, the pelvis was short and broad but it lacked some of the refinements that allow modern humans to swing their hips and walk with a striding gait. The best evidence for the morphology of the Au. afarensis pelvis comes from AL 288-1. The ilium is short, broad and has tilted backwards (‘sagittally rotated’) along with the sacrum. It too is short _-_Fig._183.gif) and broad compared to that of an ape, consisting of only five fused vertebrae rather than six, and its rotation would have brought the spine into a more upright position and an improved distribution of body weight on the vertical axis. This was not only very helpful for standing upright but also affected the operation of the deep gluteal muscles used for locomotion.
and broad compared to that of an ape, consisting of only five fused vertebrae rather than six, and its rotation would have brought the spine into a more upright position and an improved distribution of body weight on the vertical axis. This was not only very helpful for standing upright but also affected the operation of the deep gluteal muscles used for locomotion.
Femur: A number of examples of Au. afarensis femora have been recovered—most of them from Hadar but individual ones have been found at Laetoli, Maka and Dikika (DIK-1-1). They display a number of derived features that are consistent with 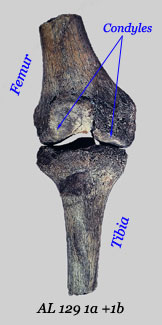 the sort of striding bipedalism that characterizes humans. In all age groups the bicondylar angle indicates a ‘valgus’ knee, which is most apparent in the first fossil to be discovered, AL 129 1a+1b. This indicates that the foot was located under the body’s centre of gravity during the walking cycle. The condyles themselves are flatter and more elliptical in shape than those found in apes, which means that the ligaments at the side of the knee tighten when the leg is fully extended. Also evident in the various specimens is an elongated femoral neck and a prominent intertrochanteric line where the iliofemoral ligament is attached. The last is the largest ligament in the body and prevents it from tipping backwards while in the standing position. All of these changes enabled hominins to walk in our characteristic ‘striding’ manner rather than the lurching style of the apes.
the sort of striding bipedalism that characterizes humans. In all age groups the bicondylar angle indicates a ‘valgus’ knee, which is most apparent in the first fossil to be discovered, AL 129 1a+1b. This indicates that the foot was located under the body’s centre of gravity during the walking cycle. The condyles themselves are flatter and more elliptical in shape than those found in apes, which means that the ligaments at the side of the knee tighten when the leg is fully extended. Also evident in the various specimens is an elongated femoral neck and a prominent intertrochanteric line where the iliofemoral ligament is attached. The last is the largest ligament in the body and prevents it from tipping backwards while in the standing position. All of these changes enabled hominins to walk in our characteristic ‘striding’ manner rather than the lurching style of the apes.
Tibia & Fibula: The tibia (shin bone) is known from incomplete fragments (AL 288-1; DIK-1-1) and a whole example from KSD-VP-1/1. It includes a number of derived modifications that suggest bipedalism. The shaft is broader at the upper (proximal) end than that of an ape. At the other end, the articular surface of the talus is perpendicular to the long axis of the shaft, unlike the great apes where it meets the talus at an angle. That would imply that, while we could point our toes up (dorsiflexion) or downwards (plantarflexion) it was largely restricted to motion back and forth on the sagittal plane. Apes have more flexibility for movement from side to side.
The fibula is represented by a number of partial specimens from the Hadar region, including both ends of the right fibula of AL 288-1, making it possible to reconstruct her ankle joint. The evidence has been interpreted as showing a greater range of plantarflexion than found in modern humans, possibly reflecting their arboreal lifestyle. Or perhaps it was simply a holdover (plesiomorphic) trait, soon to disappear.
Feet: There are a couple of dozen fossilized foot bones in the sample from AL 333 (although none in articulation) and they show a number of features consistent with bipedal locomotion. Apes have prehensile feet which operate much like a hand with an opposable thumb while hominin feet are entirely dedicated to weight-bearing. This involved wholesale changes in the arrangement of the numerous small bones that make up their structure. For example, the trochlea of the talus (the upper surface of the ankle bone, aka. talus/astragalus) is larger and less arched than is normal in apes.
The heel is the first part of the foot to hit the ground when striding forward and has to bear most of the shock when the foot hits the ground therefore the heel bone (calcaneus) of Au. afarensis is longer at the back just as it is in modern humans. The metatarsal heads, that make up the ‘balls’ of our feet, are also larger and the surface that articulates with the toes (phalanges) is broader and more domed, which means it is more conducive to dorsiflexion than plantarflexion—i.e. it is much better suited to striding than grasping.
_-_Fig.gif)
Bones of the foot of H. sapiens. Gerrish's Text Book of Anatomy
Another feature of the human foot is the longitudinal arching of the sole, which puts the spring in our step. This is accomplished by a stiffening of tarsal bones (the cuboid, cuneiforms and navicular) and is connected to the metatarsals by a ligament, known as the plantar aponeurosis, which supports the whole structure and prevents it from collapse. Apes have what is called a midtarsal break so that the foot bends at the rear (distal end) of the metatarsals rather than at the heads. Their feet are more flexible but not very well adapted for efficient bipedal locomotion over any sort of distance.
The phalanges (toes) are relatively short compared to apes and the first phalanx (also known as the hallux or big toe) was ‘adducted’, that is to say that it was lined up alongside the rest of the toes and not angled off to the side as it was among apes. This is suggested by the fossil record, in which the hallux is quite robust and has an enlarged head, and is seemingly confirmed by the Laetoli footprints (assuming they were made by Au. afarensis). The big toe in particular takes on most of the load transfer to propel the leg forward when striding forward. However, like the finger bones, the toes retain a slight, ape-like curve.
The discovery of a set of foot bones at Woranso Mille in the Afar region in 2012 (classification as yet undetermined, although clearly hominin) has raised quite a few issues. Known as the Burtele Foot, its hallux is not adducted but runs off at an angle to the other toes, indicating that it is prehensile and definitely would have altered the gait of the individual. Whatever species it belonged to, it was certainly not A. afarensis but some other hominin species. It is discussed in more detail below.
The Laetoli Footprints
These fossil, human-like footprints were discovered in 1976 by a team led by Mary Leakey at Laetoli in Tanzania. They appeared among thousands of other tracks made by animals that had moved across an ash spread laid down some 3.66 million years ago (according to the biostratigraphy and potassium-argon dating). There are a number of volcanoes, active and extinct, nearby but which one was responsible is unknown.
It must have begun to rain almost immediately, a fairly light shower that dampened the ash sufficiently to take an impression but was not hard enough to obliterate it. Traces of individual droplets could be still be seen when the site was first investigated. Apparently, the sun soon came out and dried the damp ash, which hardened like plaster around the prints. The volcano continued to throw out ash until eventually a layer up to 20 centimetres thick blanketed both the ground and the footprints. Over the millions of years that followed, additional sediments were deposited and eroded away by wind and water. Eventually the footsteps were uncovered and, by pure good fortune, were spotted by the Leakey team. Otherwise, they would have been removed by weathering within a few years.
The initial discovery (Area G) consisted of about 70 footprints extending in two lines about 25 metres long. Initially it was thought that the prints were made by two individuals walking side-by-side, either  an adult and a small child or perhaps an adult male and an adult female if one assumes significant sexual dimorphism. But the larger set of prints were somewhat ill-defined and, on closer examination it was discovered that actually belonged to a pair of adults, one following the other and stepping into its tracks. In 2015, footprints belonging to two other individuals were found in a different part of the site, Area S. No imprints of fists were found, so it is pretty clear that whoever made the tracks was not a ‘knuckle walker’ but adopted a fully upright posture. Fossil bones from Au. afarensis have been found nearby, so it is widely assumed that it was they left the tracks—the size and spacing of the tracks is consistent. However, there are plenty who disagree.
an adult and a small child or perhaps an adult male and an adult female if one assumes significant sexual dimorphism. But the larger set of prints were somewhat ill-defined and, on closer examination it was discovered that actually belonged to a pair of adults, one following the other and stepping into its tracks. In 2015, footprints belonging to two other individuals were found in a different part of the site, Area S. No imprints of fists were found, so it is pretty clear that whoever made the tracks was not a ‘knuckle walker’ but adopted a fully upright posture. Fossil bones from Au. afarensis have been found nearby, so it is widely assumed that it was they left the tracks—the size and spacing of the tracks is consistent. However, there are plenty who disagree.
Five individuals have been identified—three in Area G and two in Area S. Their estimated size has been based on the depth and length of the prints. S1 appears to have been about 165 cm tall (5’5”) and probably weighed something like 45 kg (100 lbs). This is considerably taller and heavier than any of the others and suggests it was probably a male. The others, who are estimated to range in height from 114-145 cm (3’9” to 4’9”) and may well have been females or, in the case of the smallest example, a juvenile.
These tracks represent the earliest direct evidence of bipedalism and clearly shows that it had appeared several hundred thousand years before tool use and large brains—originally these were thought to have arrived as a package marking the arrival of ‘humanity’. The fossil footprints are very similar to the ones we leave walking barefoot on a wet, sandy beach. Their feet were arched and the hallux was adducted, like ours, alongside the rest of the toes rather than in opposition to them, as with apes. When they walked, the heel hit the ground first followed by a weight transfer to the ball of the foot before pushing off again with the toes.
Contemporaries
Australopithecus bahrelghazali: In 1995, what is claimed to be a new hominin species represented by three mandible fragments and an isolated tooth (KT-12/H1), was identified at a site called Koro Toro in the Sahel region of Chad, a region lying some 2,500 km west of the Rift Valley. The  sediments associated with the fossils have been dated to about 3.5 million BP and are therefore contemporary with Au. afarensis. They also indicate a widespread distribution for australopithecines at this time. The Sahel was not so arid as today and would be characterized as open savannah— grasslands with some tree cover. There were a number of large herbivores around including Proboscidea, Rhinocerotidae and Giraffidae along with smaller ones such as hipparions
sediments associated with the fossils have been dated to about 3.5 million BP and are therefore contemporary with Au. afarensis. They also indicate a widespread distribution for australopithecines at this time. The Sahel was not so arid as today and would be characterized as open savannah— grasslands with some tree cover. There were a number of large herbivores around including Proboscidea, Rhinocerotidae and Giraffidae along with smaller ones such as hipparions
An analysis of carbon isotopes in their teeth suggest that they lived mainly on a diet of C4 foods—grasses, tubers and rhizomes etc.—as well as some fruit, leaves, pods, and insects. The dental array of the two species is very similar. The ‘alveolar process’, where the sockets for the incisors are located, is angled differently and the premolar has three roots rather than two but most scholars think these are insufficient grounds for separating the two species.
Kenyanthropus platyops: In 1998 and 1999 an expedition led by Maeve Leakey and her daughter Louise collected another set if anomalous fossils in a survey of the Lomekwi region on the west shore of Lake Turkana in Kenya. The finds include two partial maxillae, a complete temporal bone (which houses the structure of the ear), some isolated teeth and, crucially, a largely complete if badly distorted cranium (KNM-WT 40000) which became the holotype of a new species they named Kenyanthropus platyops (“flat-faced man from Kenya”) in 2001. During the course of these two seasons, no less than 34 skull fragments were found—although it is uncertain how many of them belong to the same species. The fossils all come from a deposit that was formed between 3.57 and 3.4 million BP which places it securely within the A. afarensis time frame.
Despite the distorted condition of the cranium which had splintered into over a thousand fragments, a number of conclusions could be drawn about its morphology. In many respects it resembles later species such as H. habilis and H. rudolfensis. The most obvious feature was, as the name implies, its flat face which is evident by the reduced slope of the area between the nose and the mouth (the nasoalveolar clivus) compared to other australopithecines. The zygomatic arches (cheekbones) are taller, steeper and start much further forward on the face; and the nasal opening is narrower. The face lacks the anterior pillars, an externally prominent bony column running vertically alongside the nasal aperture, on Au. afarensis. The molars are much smaller than those of A. afarensis or any other early hominin. It was not possible to measure the cranial capacity of the skull but there is no reason to think that it is not much different than that of contemporary hominins.
In 2011 a substantial collection of stone tools was found in the Lomekwi. The tools were very simple and crudely made but it is clear that they were not products of natural forces but had been deliberately shaped for a purpose. They are discussed in more detail below.
Australopithecus deyiremeda: Evidence of yet another early hominin species was discovered by a team led by Dr. Yohannes Haile-Selassie at Woranso–Mille in the central Afar region of Ethiopia in -and-BRT-VP-3-14-shown-in-occlusion.png) 2011. The initial find and holotype (BRT-VP-3/1) was a partial maxilla with most of its teeth, but two fossil mandibles one complete, (BRT-VP-3/14) and one broken (WYT-VP-2/10) were found not far away (the paratypes). All of them were surface finds weathered out of sandstone deposits that have been dated to between 3.3 and 3.5 million BP, well within the time frame of A. afarensis.
2011. The initial find and holotype (BRT-VP-3/1) was a partial maxilla with most of its teeth, but two fossil mandibles one complete, (BRT-VP-3/14) and one broken (WYT-VP-2/10) were found not far away (the paratypes). All of them were surface finds weathered out of sandstone deposits that have been dated to between 3.3 and 3.5 million BP, well within the time frame of A. afarensis.
An analysis of the fossils shows that, while they are pretty obviously australopithecine, there are differences between them and any of the other species found to date. As was the case with most australopithecines the jaw was prognathic, but its robustness is more similar to the genus Paranthropus if somewhat smaller. The molars and premolars are smaller than those of A. afarensis, but with relatively thick enamel. These adaptations suggest a diet that involved a lot of hard chewing—masticating leaves and crunching nuts, for example. What all of this would seem to indicate is that there were two australopithecine species operating in the same region at the same time but exploiting different elements of the ecosystem.
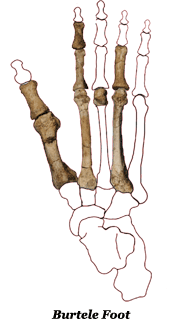 The Burtele Foot: The following season, 2012, saw the rare discovery a partial foot by Dr. Haile-Selassie’s team not far from the previous year’s finds at Woranso-Mille. It is not directly associated with them but the context falls into the same time frame, c. 3.5 million BP. The fossil (BRT-VP-2) includes most of the right forefoot, the metatarsals and phalanges. Essentially it had a mix of features, some derived from the earlier hominin Ardipithecus ramidus, who lived about a million years earlier (c. 4.5 million BP), and others that it shares with Australopithecus afarensis. The tarsals and phalanges of the outer toes are sufficiently arched and their heads are domed and angled to put a bit of a spring in the step and make fully bipedal walking possible, if somewhat awkward. What really distinguished it from contemporary australopithecines was its abducted hallux (its prehensile big toe), similar to that of Ardipithecus. This was a very useful feature if climbing trees was a regular part of your existence but not if you were spending most of your time out in the open. A lot of propulsive power is generated by the big toe.
The Burtele Foot: The following season, 2012, saw the rare discovery a partial foot by Dr. Haile-Selassie’s team not far from the previous year’s finds at Woranso-Mille. It is not directly associated with them but the context falls into the same time frame, c. 3.5 million BP. The fossil (BRT-VP-2) includes most of the right forefoot, the metatarsals and phalanges. Essentially it had a mix of features, some derived from the earlier hominin Ardipithecus ramidus, who lived about a million years earlier (c. 4.5 million BP), and others that it shares with Australopithecus afarensis. The tarsals and phalanges of the outer toes are sufficiently arched and their heads are domed and angled to put a bit of a spring in the step and make fully bipedal walking possible, if somewhat awkward. What really distinguished it from contemporary australopithecines was its abducted hallux (its prehensile big toe), similar to that of Ardipithecus. This was a very useful feature if climbing trees was a regular part of your existence but not if you were spending most of your time out in the open. A lot of propulsive power is generated by the big toe.
Given that the Burtele foot does not belong to Au. afarensis nor to whoever was responsible for the Laetoli footprints, and given the fact that it was found in close proximity to the fossil remains of Au. deyiremeda, and that the foot suggests a more arboreal lifestyle, the likeliest candidate for ownership would seem to be the latter.
Summary: The operating assumption at the end of the last millennium was that there ought to be a clear line of descent from ape to man, and it was argued that there was a single strand of evolution and that A. afarensis was the common ancestor of all later hominin species. However, the more we have learned since then, the muddier the waters seem to get. It now seems credible that there were at least four species of australopithecine operating in the Rift Valley System in the Sahel. The differences in the cranial morphologies of Au. afarensis, Au. bahrelghazali, Au. deyiremeda, and K. platyops are sufficiently distinct as to warrant that assessment—although there are plenty of palaeoanthropologists who would disagree. Complicating the established picture is clear evidence from the Human Genome Project that species interbreeding took place during the Middle Palaeolithic in Eurasia between members of the genus Homo.
Lifestyle
Judging from the faunal remains recovered from the region, the landscape was characterized by open grassland and scrub (savannah) with stands of isolated woodland and solitary trees. Denser forest areas were located in river valleys and lake margins. Although a bit more plentiful than today, rainfall was seasonal and there were long dry spells when food was scarce. The woodlands were dominated by plants that photosynthesized using C3 carbon fixation, which made them very susceptible to heat and aridity. The C4 grasses of the savannah were less nutritious but much hardier, using much less water to carry out photosynthesis, and were better able to withstand the hot, dry conditions. So, when the dry season came, the animals with the best chance of survival were those who were able to exploit those resources. The most prominent among them were the large, herd grazers whose digestive systems were capable of processing the immense quantities required but many browsing animals that normally ate mainly leaves and fruit from the trees could cope with grass if necessary. Along with the herbivores there came a number of large carnivores, big cats and hyenas, who fed on their flesh. Studies of the ratio of carbon isotopes in the bones and teeth can determine how much of each an animal consumed.

Savannah in the Taita Hills of Kenya
This evidence suggests australopithecines of this time and place were omnivorous feeders, consuming whatever was to hand. C3 fruits and leaves from the wooded areas made up most of their diet but they also consumed, directly or indirectly, C4 plants such as grasses, found out in the savannah, as well as sedges found mainly in wetlands. While there is no solid evidence for scavenging animal flesh, it is generally believed that this must have taken place. Such wide ranging food procurement indicates that Au. afarensis exploited a range of different habitats and was capable of operating in each. In order to do so successfully in the open plain, it had to be able to cover many times the amount territory as in the forest where food (when in season) could be found with little effort. The method of locomotion they adopted was bipedalism which enabled it to move at a reasonable pace for distances of several kilometres. By raising its head above the grass, it was able to spot potential danger in time to take countermeasures—especially if there were multiple pairs of eyes on the lookout. It also freed the hands to carry food to a safer place to share it out or to carry helpless infants. The fact that the species retained some ability to climb was also very useful when danger threatened.
Stone Tools
In 201o, fossil animal bones bearing cut marks made by stone tools were found at Dikika, a site in the Afar region of Ethiopia, and dated to some 3.3 million years ago. No hominin remains were found at the site and only Au. afarensis fossils were found in the region at this time so they are the most likely candidates. No actual tools were found at Dikika but they did turn up at Lomekwi 3 on the western shore of Lake Turkana (Kenya). Cobbles of stone can be battered by the natural forces of wind, water and gravity, but the fracture patterns do not suggest that that was the case here. Chimpanzees are well-known to have bashed stones together using a ‘hammer and anvil’ technique to break open nuts, creating fracture patterns. The Lomekwi finds, however primitive, give every impression of having been deliberated flaked and shaped, rather than simply bashed about by river action or having been randomly used in percussive activities.
Nearly 150 pieces were recovered of which the majority were cores (83) or flakes (35). Of the rest, there were some that may have been used as hammers with others possibly served as anvils. The stone, predominantly basalts and phonolites, probably all came from a conglomerate located only 100 metres away and none of it had moved very far in the last 3½ million years. The cores are large and hefty compared to the much later samples from Olduvai Gorge, and average a little over 3 kilograms. Flakes had a wider range in size (c. 20-200 mm) and were much thinner on average. Anvils were very large and heavy, 30 kilos or more, and the upper surface was generally flat. The flakes were struck off the core using either the ‘passive hammer’ technique, whereby they were bashed against a heavy stone anvil, or the bipolar method, which involved placing the core on the anvil and then striking the opposite end with a hammer. They are classified as Pre-Mode 1 according to the system devised by Grahame Clark.
The tool-makers were not the most adept stone workers and there was a lot of wastage, but it is clear that there was some planning and foresight involved as well as a certain amount of manual dexterity. The Lomekwi finds show that these characteristics were already present well before Homo habilis and the Oldowan industry.
Further Reading
| Aronson, J. & M. Taieb | (1981) | Geology and Paleogeography of the Hadar Hominid Site, Ethiopia. In: Rapp G & Vondra CF, editors. Hominid Sites: Their Geologic Settings |
| Ayala, F. J. and C. J. Cela-Conde | (2017) | Processes in Human Evolution. Second Edition |
| Barham, L. & P. Mitchell | (2008) | The First Africans. African Archaeology from the Earliest Toolmakers to Most Recent Foragers |
| Bonnefille, R. et al | (1987) | Palynology, stratigraphy and palaeoenvironment of a Pliocene hominid site (2.9-3.3 MY) at Hadar, Ethiopia. Palaeogeography, Palaeoclimatology, Palaeoecology. 60 |
| Cartmill, M. & F.H. Smith | (2022) | The Human Lineage. Second Edition |
| Clark, J. Grahame | (1969) | World Prehistory: A New Outline |
| Haile-Selassie, Y. et al | (2012) | A new hominin foot from Ethiopia shows multiple Pliocene bipedal adaptations. Nature 483 |
| (2015) | New species from Ethiopia further expands Middle Pliocene hominin diversity. Nature 521 | |
| Harmond, S. et al | (2015) | 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature 521. |
| Harrison, T. | (2011) | “Laetoli Revisited: Renewed Paleontological and Geological Investigations at Localities on the Eyasi Plateau in Northern Tanzania”. In Paleontology and Geology of Laetoli: Human Evolution in Context: Volume 1. Edit. Terry Harrison |
| Johanson, D. & Maitland Eddy | (1981) | Lucy. The Beginnings of Humankind |
| Kimbel, William H., & Lucas K. Delezene | (2009) | “Lucy Redux: A Review of Research on Australopithecus afarensis”. Yearbook of Physical Anthropology. Institute of Human Origins, Arizona State University |
| Leakey, Mary D. & J.M. Harris eds. | (1987) | Laetoli: A Pliocene Site in Northern Tanzania. |
| Leakey, Maeve, F. Spoor, et al | (2001) | New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature 410 |
| Lewen, Roger | (2005) | Human Evolution. 5th Edition. |
| Maslin, M | (2017) | The Cradle of Humanity |
| Maslin, M., et. al. | (2014) | East African climate pulses and early human evolution. Quaternary Science Review. |
| Owen-Smith, N. | (2021) | Only in Africa. The Ecology of Human Evolution |
| Rossouw L., & L. Scott | (2011) | Phytoliths and Pollen, the Microscopic Plant Remains in Pliocene Volcanic Sediments Around Laetoli, Tanzania. Paleontology and Geology of Laetoli: Human Evolution in Context. |
| Scarre, C. (edit.) | (2018) | The Human Past. Fourth Edition |
| Su, D.F. & T. Harrison | (2007) | “The paleoecology of the Upper Laetolil Beds at Laetoli”. in Hominin Environments in the East African Pliocene: An Assessment of the Faunal Evidence. |
| Zaitsev, A.N. et al | (2023) | Laetoli: The Oldest Known Hominin Footprints in Volcanic Ash. Elements |
