



_male_with_cattle_egret.jpg)
.png)
.png)





.jpg)


The Pleistocene (c. 2.58 million-12 thousand BP)
The Pleistocene Epoch marks the beginning of what we call the Ice Age (although it only the latest in a series of ‘ice ages’ that goes back at least 2.4 billion years). In this instance, it was triggered by the collision of North and South America which formed the Isthmus of Panama, blocking the flow of warm water from the Pacific into the Atlantic Ocean. Instead, cold water from the polar regions spread into the Atlantic leading to a sharp drop in annual temperature throughout the planet and the formation of glacial ice in the northern hemisphere. During this time, the climate of the Eurasian continent and North America was characterized by a continuous cycle of advancing and retreating ice sheets (glacials and interglacials). At least six of these have been identified during the Pleistocene.
At the beginning of the Pleistocene, the glacial cycles averaged about 41,000 years but, about a million years ago, they became much longer, lasting for about 100 thousand years or so. Interglacials during this period were much shorter, lasting anywhere from 10 to 30,000 years. The effects the prolonged cold were dramatic. The ice sheets covering North America and Greenland could be as much as 3-4 kilometres thick and advanced as far south as 40 degrees latitude. The amount of water turned into ice resulted in a drop in global sea level by as much as 120 metres. The flow of ocean currents and the direction of the prevailing winds were affected, changing weather patterns all over the globe. The frigid temperatures meant less moisture was evaporated from the world’s oceans and therefore less was available to fall as rain.

Note the magnitude of the changes post-750 thousand BP
The extreme cold that characterized the northern hemisphere was much less pronounced further south where the problem was one of increased aridity and its impact on the local ecosystems. There were fluctuations of course but even so the long term trend was to increased aridity and cooler temperatures. Dry seasons became more extreme and might span several months putting an enormous strain on the ecosystem as vegetation withers away. The vast rainforests that covered most of the land became steadily reduced while areas of grassland expanded. Obviously, this had an impact on the animal life. In good years, when forage is abundant the number of herbivores can be vast, but given a long dry season or a series of them over a number of years, there would have been a large die-off, especially among the more vulnerable—the very young and the very old. The long-term effects of climate change can be seen in the faunal record. At the beginning of the Pleistocene, browsers made up the bulk of the herbivore populations with grazers amounting to 15-25% of the total but the rapid increase in aridity about 1.8 million years ago saw the proportions become nearly equal.
A wide range of grazing animals could be found in East Africa according to the fossil record. These included a number of species that have long since become extinct. There was a giant wildebeest (Megalotragus), a giant buffalo (Syncerus antiquus), which stood about 2 metres tall at the withers and weighed as much as 4 tonnes, and a giant warthog (Metridiochoerus)— a formidable beast, double the size of the modern version and with two pairs 0f large tusks. The situation is broadly similar in South .jpg) Africa where a large number of fossils have been recovered from caves at sites such as Swartkrans,Sterkfontein and Makapansgat. Reduncine grazers (reedbucks and waterbucks, for example)—species that are normally associated with watery environments—are much fewer in number.
Africa where a large number of fossils have been recovered from caves at sites such as Swartkrans,Sterkfontein and Makapansgat. Reduncine grazers (reedbucks and waterbucks, for example)—species that are normally associated with watery environments—are much fewer in number.
More modern species begin to assert themselves as the Pleistocene continues. Alcelaphine antelopes and gazelles, including the hartebeest and the topi, show up in the fossil record (thin though it may be) in increasing numbers after 900,000 BP. They and the remaining grazers appear to have turned their attention from C3 to the more drought-resistant C4 grasses although they remained vulnerable during the dry season. Many of the largest Pliocene grazers disappeared from the record in both the Rift Valley and South Africa at this time. These included Reck’s elephant, the short-necked giraffe, the all of the others mentioned above.
A similar fate met the larger predators—the sabre-tooths and the giant hyaena—which had all vanished from the scene by the end of the Pleistocene. These were typically ambush hunters which lurked in the rapidly disappearing forests. Moving up the food chain were members of the genus Panthera, including lions, leopards and cheetahs, which first appeared late in the Pliocene (ca. 3.5 million BP) along with modern species of hyaena and the first canids—wild dogs, jackals and foxes. All of them relied mainly on speed and chased down their prey in the open.

Olduvai Gorge
The Palaeolithic
The Pleistocene also saw the arrival of a new predator on the scene, one that would in time achieve dominance— Man (Homo). His presence is indicated, not only by fossilized bones, but by the presence of the stone tools that he made and used to expand his physical powers. Their appearance marked the beginning of what is known as the Palaeolithic (in Europe) or Old Stone Age (in Africa) which lasted over 2.5 million years—by far the longest stretch of time in human history.
Homo habilis
Up until quite recently it was generally believed that one of the defining characteristics of being human was the manufacture and use of tools. So in 1960, when Louis Leakey’s team working at Olduvai Gorge (Tanzania) discovered a novel set of fossil hominin remains in the same deposits where simple stone artefacts had been found in the 1930s. Although not directly associated with the objects, he was quick to classify the remains as human and named the new species Homo habilis (“handy man”). However, in the decades that followed, evidence for stone working has been found in contexts where humans are absent but australopithecines are present. The problem has been made even more complex by the fact that craniodental remains (i.e. skulls and teeth) are rarely found in association with those of the rest of the body and, when they are, they are generally quite fragmentary. Under the circumstances, the matter of the classification of the new species, Homo or Australopithecus, remains open to debate.  Further complicating the picture is the fact that there is a wide variation in the morphology of the various fossils included in the designation. This fact has been attributed to sexual dimorphism by some anthropologists but others think that it may reflect similar but different species (H. rudolfensis, see below). This debate will be explored in more detail in a future article.
Further complicating the picture is the fact that there is a wide variation in the morphology of the various fossils included in the designation. This fact has been attributed to sexual dimorphism by some anthropologists but others think that it may reflect similar but different species (H. rudolfensis, see below). This debate will be explored in more detail in a future article.
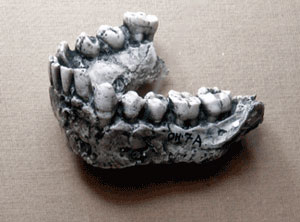
As far as the timeline is concerned, the initial discovery (OH7) was dated to about 1.75million BP but subsequent finds have pushed the date date back as much as 750,000 years, to the beginning of the Pleistocene. The first stage of the Pleistocene is known at the Gelasian (c. 2.58-1.8 million BP) and coincided with the initial advance of the ice sheets in the northern hemisphere and the onset of increased aridity in Africa. This led to an expansion of the savannah at the expense of the rainforest.
H. habilis was taller (100-148 cm) and heavier than most australopithecines, with a larger brain (600–650 cc vs. 380-500cc), longer legs and slightly flatter faces. Their feet are reasonably well-suited for bipedal locomotion but, with elongated toes and flexible ankles, they were adept climbers as well. The latter was important because they still relied to a large extent on that ability to access fruits and the more succulent leaves. It also gave them access to relatively secure places to spend the night.
What really marks them out from their predecessors was their dentition. Their molars were smaller with thinner enamel, which may signify a shift from tough, fibrous plant food to a more wide ranging diet that included more animal protein. Isotope analysis of their tooth enamel indicate a diet made up of 25-50% C4 plant material, much of which probably came from consuming the scavenged remains of grazing animals. As far as the consumption of meat is concerned, cut marks made by stone tools generally overlay the tooth marks of carnivores on the bones of larger animals, indicating that they were scavenged, but the opposite is the case on many of the smaller mammals, which suggests that they had been killed through hunting.
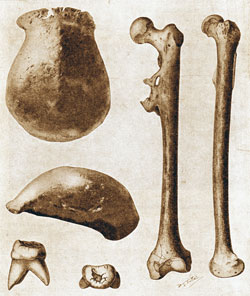
Homo rudolfensis: In 1972 a somewhat larger hominid skull (KNM-ER 1470) was discovered by Richard Leakey beside Lake Turkana (then known as Lake Rudolf) in Kenya in deposits dating to 2.1-1.95 million years ago. Apart from its size, which could be explained by sexual dimorphism in the H. habilis species, there were distinctions in its dentition and the shape of the skull. Its cranial capacity was estimated by Leakey at ca. 800 cc but this has been disputed. Unfortunately, although postcranial elements were found nearby, none were found in association with skulls or skull fragments. The big unanswered question is whether H. rudolfensis was a distinct contemporary of H. habilis or if it was a chronospecies (i.e. one that evolved in a direct line from the latter through natural selection).
The Oldowan Industry (Mode 1)
The stone tools themselves belong to the Oldowan tradition which is classified as Mode 1, which means they consist mainly of pebble cores and the flakes struck from them. Evidence (obtained through  refitting of flakes) from Lokalalei 2C at West Turkana in Kenya (Delagnes & Roche, 2005) where nearly 3,000 artefacts were found, suggests that there was a strategy for shaping tools, however primitive. The cores were selected for quality and the flakes were struck in a controlled manner indicating a degree of manual dexterity. Evidently the tool makers understood the physical properties of the raw material and the mechanics of flake production. The cores might be pointed or edged and are often described as ‘choppers’ but may have been used for a variety of purposes— digging up roots and tubers or cracking open bones, for example. However, it is also quite possible that they represent the discarded by-products of flake production. The flakes would have then be used for scraping bones and slicing meat.
refitting of flakes) from Lokalalei 2C at West Turkana in Kenya (Delagnes & Roche, 2005) where nearly 3,000 artefacts were found, suggests that there was a strategy for shaping tools, however primitive. The cores were selected for quality and the flakes were struck in a controlled manner indicating a degree of manual dexterity. Evidently the tool makers understood the physical properties of the raw material and the mechanics of flake production. The cores might be pointed or edged and are often described as ‘choppers’ but may have been used for a variety of purposes— digging up roots and tubers or cracking open bones, for example. However, it is also quite possible that they represent the discarded by-products of flake production. The flakes would have then be used for scraping bones and slicing meat.
The industry has usually been associated with H. habilis but new evidence suggests that very similar tools first appeared about 2.9 million years ago on the Homa Peninsula near the shores of Lake Victoria in Kenya. This would be about 500,000 years before the first recorded appearance of the first humans, and it would appear that the tools were actually made by Australopithecus garhi, the only hominin whose remains have been found in the area at that time.
Paranthropus
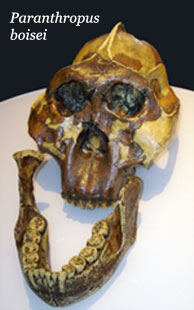 At about the same time as H. habilis emerged, another genus of hominins known as Paranthropus appeared in East Africa. The first to be identified was P. aethiopicus (2.6 million BP) whose fossils were found in Ethiopia, but other species appeared subsequently, P. boisei (2.3 million BP) in Tanzania and P. robustus (2 million BP) in S0uth Africa. They were all rather slight in stature and are distinguished from the more ‘gracile’ australopithecines (from whom they descend) by their massive skulls and powerful, ‘nutcracker’, jaws. Carbon isotope and microwear analyses indicate that they lived primarily on C3 forest plants and C4 savannah plants suggesting that they moved between the two areas according to the season, consuming fruit, nuts and leaves in the former and underground resources, such as roots, tubers and rhizomes in the latter.
At about the same time as H. habilis emerged, another genus of hominins known as Paranthropus appeared in East Africa. The first to be identified was P. aethiopicus (2.6 million BP) whose fossils were found in Ethiopia, but other species appeared subsequently, P. boisei (2.3 million BP) in Tanzania and P. robustus (2 million BP) in S0uth Africa. They were all rather slight in stature and are distinguished from the more ‘gracile’ australopithecines (from whom they descend) by their massive skulls and powerful, ‘nutcracker’, jaws. Carbon isotope and microwear analyses indicate that they lived primarily on C3 forest plants and C4 savannah plants suggesting that they moved between the two areas according to the season, consuming fruit, nuts and leaves in the former and underground resources, such as roots, tubers and rhizomes in the latter.
They had particularly large molars with thick enamel which microwear analysis suggests were used to deal with the hard seeds, tubers and corms that made up their ‘fallback foods’. Their peculiar dentalwork may well have obviated need the for stone tools, even though it has been suggested that they may have been responsible for at least some of Mode 1 assemblages as their fossils have been found in contemporary deposits nearby. Ultimately, a predominantly plant based diet proved inadequate for their survival and the species died out about 1.4 million BP.
Homo erectus/ergaster
At some point towards the end of the Gelasian stage (ca. 2.58-1.8 million BP), a new hominin species appeared in Africa, Homo ergaster. Its origins are somewhat murky and it turns up in East Africa quite suddenly. It coexisted with H. habilis for several hundred thousand years but, given its sudden appearance, does not appear to be an offshoot of that particular branch of the human family tree. More importantly, a very closely related if not identical species (the debate rages on) appeared in Eurasia at roughly the same time. The usual explanation is that this is evidence of a migration—the first such among hominins—out of Africa. But, since priority has not been established, the possibility remains that the migration may have gone the other way.
The first discoveries were made early in the last century in East Asia, in Indonesia and China. Known popularly as ‘Java Man’ and‘Peking Man’ they came to be classified as Homo erectus (‘upright man’) and have been dated to somewhere around 700 thousand years BP. The earliest Homo ergaster fossil recovered to date is part of the cranium, an occipital bone belonging to an infant that was found in the Drimolen Palaeocave System near Johannesburg, South Africa and dating to ca. 2.04-1.95 million BP. However, the attribution is uncertain. Otherwise, most of the African fossils come from the vicinity of Lake Turkana in Kenya and fall into the date range 1.8-1.7 million BP).
The big question at the moment is whether or not they are separate species—one characteristic of Africa and the other, Asia—or if one is simply a variant of the other given the distances, both in time and in space, that separate the various fossils. The most important physical differences between the two are to be found in the shape of the skull and their facial features—the presence or absence of ‘keeling’ (ridges) along the top and sides of the skull and the configuration of the brow ridge for example. But are these distinctions great enough to warrant classification into separate species?
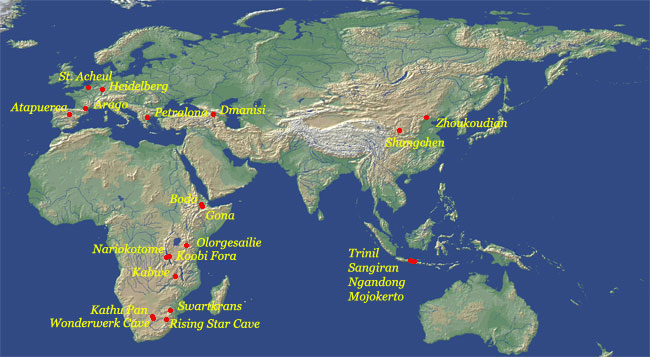
Distribution of sites associated with Homo Erectus and related species (click map for larger version)
Fossils belonging to Homo ergaster can be distinguished from those of H. habilis on the basis of their size, the morphology of the skull and its dentition. H. ergaster was bigger and taller (ranging from 146-185 cm. in height (well within the range of modern humans)— and had a much larger brain (900–1,200 cc). These developments came at a cost, however, as large, big-brained offspring took much longer to grow and mature, and required a higher caloric intake while they were doing so. Brain tissue, for its weight, requires nine times the energy intake as the rest of the body. The most abundant and obvious source for 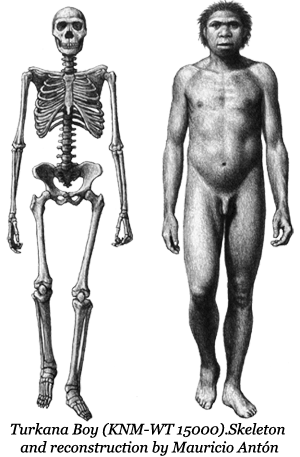 the additional fats and proteins needed were the medium to large sized ungulates that grazed the savannah. Up until now it had been possible to acquire these by scavenging the kills of larger, more adept predators but, with the greater amounts required, it was much more efficient and reliable to kill these animals yourself.
the additional fats and proteins needed were the medium to large sized ungulates that grazed the savannah. Up until now it had been possible to acquire these by scavenging the kills of larger, more adept predators but, with the greater amounts required, it was much more efficient and reliable to kill these animals yourself.
With their long legs, stiff ankles and narrow hips, they were able to develop a more efficient gait that enabled them to cover long distances at quite a respectable pace. Other anatomical changes helped give them the additional stamina required. The bridge of the nose is high and prominent which helped them conserve moisture when exhaling— very helpful in an arid environment. In addition, although there is no direct evidence, it seems likely that it was probably about this time that humans lost most of their thick body hair. This facilitated sweating, reducing the chances of overheating while in hot pursuit of game—especially under the hot midday sun when the grazers were at their weakest and most vulnerable and when their main rivals (especially the big cats) tended to rest and snooze. Although faster over short distances, the grazers out on the savannah did not have the same level of stamina as their human pursuers having to stop, sooner or later, when heat exhaustion set in. Then they could be dispatched with relative ease. It seems likely that these early hunters used wooden spears to dispatch their prey— either by stabbing them or launching missiles at them. Changes in the configuration of their shoulders would have enabled the latter. However, hunting is a very chancy business and a large part (probably the largest part) of their diet was provided by plant resources—fruits and leaves along with underground storage organs such as roots, tubers and rhizomes. The enamel on their teeth was much thinner which made them less suited for crunching hard foods like nuts but better for shredding and masticating the tougher ones that they now came to on more and more.
The earliest evidence of H. ergaster, physical and cultural, comes from the area surrounding Lake Turkana in Kenya. Initially, they seem to have confined their activities to the edge of the forest that enveloped the lake and flanked the river valleys that led away from it where they found fresh water and secure refuge—much the same territory that H. habilis exploited— but soon they began range well out into the savannah and beyond. Being significantly larger than their predecessors, they needed more territory to sustain them—especially in light of climate change and their increased dependency on animal protein. It is widely believed that the impulse for these movements was a change from cool and dry conditions to warm and moist, which allowed early humans to thrive and their numbers to grow. Fresh pastures were needed and, with the amelioration of the climate, there were no longer great expanses of desert to hem them in. Within a relatively short time they could be found throughout the continent and beyond.
Out of Africa 1(or 2?)
Early humans were present in Eurasia from about 2 million years ago. Fossils have been found at Dmanisi in Georgia, dating to about 1.85 million BP—in other words, 200,000 years earlier than the earliest uncontested H. ergaster in Africa. There were 5 skulls and hundreds of postcranial bones. Although they are similar to H. ergaster in many respects, their slight build, dentition and smaller  brains were more akin to H. habilis. The stone industry is strictly Mode 1 with no bifaces or Developed Oldowan tools. It has been argued that they may be the descendants of an earlier wave of migration out of Africa and that they represent a separate species—Homo georgicus. To further complicate the picture, simple stone tools dating to about 2.1 million BP have recently been recovered at Shangchen in central China. The date is at least as early as any of the African dates for H. ergaster and lends support for the possibility of a previous migration (of ‘habilines’?) and a ‘reverse migration’ of ‘erectines’.
brains were more akin to H. habilis. The stone industry is strictly Mode 1 with no bifaces or Developed Oldowan tools. It has been argued that they may be the descendants of an earlier wave of migration out of Africa and that they represent a separate species—Homo georgicus. To further complicate the picture, simple stone tools dating to about 2.1 million BP have recently been recovered at Shangchen in central China. The date is at least as early as any of the African dates for H. ergaster and lends support for the possibility of a previous migration (of ‘habilines’?) and a ‘reverse migration’ of ‘erectines’.
The most obvious route out of Africa (and back in again) was by way of the Sinai which forms a land bridge connecting it with southwest Asia but, owing to fluctuations in sea-level, a similar connection may have also existed at the southern end of the Red Sea linking Djibouti and Yemen. On the other side of the Mediterranean, early humans had reached Italy and the Iberian Peninsula by as early as 1.3 million BP. Although an overland route via the Caucasus or Anatolia seems most likely, but crossing the Straits of Gibraltar or island hopping from Tunisia to Sicily was certainly possible.
The Acheulean Industry (Mode 2)
Not long after the arrival of H. ergaster on the scene, a very distinctive stone tool technology known as the Acheulean after Saint Acheul on the River Somme in France, where it was first identified in 1859. Despite its name, the industry originated in Africa, however, and probably derived from the Oldowan—although there is no clear evidence of that. The earliest finds so far date to about 1.7 million years ago (from  Gona in Ethiopia and by Lake Turkana in Kenya). The industry is also well-represented in Eurasia and have been found in China dating to ca. 1.5 million BP and in Europe some half a million years later.
Gona in Ethiopia and by Lake Turkana in Kenya). The industry is also well-represented in Eurasia and have been found in China dating to ca. 1.5 million BP and in Europe some half a million years later.
The products of the Acheulean are classified as Mode 2, which means that a hammer was used to strike off small flakes from an original core or large flake to produce a tool of predetermined size and shape that was lighter and better suited to its task. The diagnostic form is the ‘hand-axe’, which was knapped symmetrically and on both sides (bifacially) to produce an oval or heart-shaped product with a single edge that ran around most of the perimeter. These could then be used as cleavers, digging tools or picks. It was a much more sophisticated technique than the Oldowan, requiring a preconceived idea of what the end result would look like; a well thought out plan for the sequence of flake removal; and careful execution.
By about 750 thousand BP, made of antler, bone or wood, was used to detach even smaller flakes to produce a much finer tool. The flakes initially struck off were often “retouched” to make useful cutting and piercing tools and subsequent retouching made them even more so. Mode 2 production yielded more cutting edge from the original raw material, making their toolkit lighter and more transportable. In addition to the technological changes evident in Mode 2, there was a huge distinction in tool usage. There were more of them around and a greater variety of tool type—they were evidently much more important in people’s everyday lives.
 Although wood rarely survives from this period, it is highly probable that it was used to fashion spears and digging sticks. A wear-use analysis of points recovered from Kathu Pan, South Africa suggests that, by about 400,000 BP, many of them were hafted as spearheads (Wilkins, 2012). Long bone fragments and horn cores—sometimes shaped by grinding—have been found at Sterkfontein in South Africa and may have been used to open up termite mounds. Worked bone tools, which included a few bifaces, were recovered from Beds I and II at Olduvai Gorge. These appear to have been used for the same sorts of butchering and digging activities as the stone versions. As we have already seen, wood and bone may also have been used as percussive tools in flint working.
Although wood rarely survives from this period, it is highly probable that it was used to fashion spears and digging sticks. A wear-use analysis of points recovered from Kathu Pan, South Africa suggests that, by about 400,000 BP, many of them were hafted as spearheads (Wilkins, 2012). Long bone fragments and horn cores—sometimes shaped by grinding—have been found at Sterkfontein in South Africa and may have been used to open up termite mounds. Worked bone tools, which included a few bifaces, were recovered from Beds I and II at Olduvai Gorge. These appear to have been used for the same sorts of butchering and digging activities as the stone versions. As we have already seen, wood and bone may also have been used as percussive tools in flint working.
Use of Fire
Brush fires are a recurring feature of the African savannah—most often the result of lightning strikes that herald the arrival of the monsoons at the end of the dry season. They served a vital ecological purpose by clearing the landscape of a lot of dead growth, especially among the grasses. The regeneration that appeared with the first rains attracted large numbers of grazers along with the hunters and scavengers that depended on them, including early humans.
The benefits of fire must have been apparent early on and humans soon took advantage of them. Fire provided warmth and security from predators at night, and firelight extended the amount of time available for work or social activity. Just as important, fire had a profound effect on their diet. It would have soon become apparent that the flesh of cooked animals was much easier to chew and digest. Roots and tubers need to be broken down before they can be eaten, which can be done either through a lot of pounding or by cooking. In fact, many of them are indigestible and far less nutritious without cooking. The same applies to meat but, additionally, cooking meat got rid of most parasites, including tapeworms if it was cooked long enough. On the technological side, preheating quartzites such as flint and chert made them more brittle and gave the knapper more control over the flaking process.
cooked animals was much easier to chew and digest. Roots and tubers need to be broken down before they can be eaten, which can be done either through a lot of pounding or by cooking. In fact, many of them are indigestible and far less nutritious without cooking. The same applies to meat but, additionally, cooking meat got rid of most parasites, including tapeworms if it was cooked long enough. On the technological side, preheating quartzites such as flint and chert made them more brittle and gave the knapper more control over the flaking process.
There is no conclusive evidence of early humans actually making fire until after ca. 1 million BP, but it is highly likely that they had been exploiting it much earlier. All they required was a container made from fire-resistant leaves in which to transport glowing embers. Fire-cracked rock and burnt bone has turned up at a number of sites but so far not directly associated with humans. Patches of oxidized clay have also been found associated with cultural material, which might indicate the presence of a campfire but might equally represent the burnt stump of a tree where people had once gathered.
Lifestyle
It is estimated (based on studies of other primates) that early human groups may have numbered as many as 60-80 individuals. The members of the group would almost certainly have organized themselves on the basis of age and gender. Adult males of the species were larger and able to move faster than the females and were therefore suited to the roles of hunters and defenders of the group. Females were largely (but perhaps not entirely) precluded from the hunt because their role as child-bearers and child-rearers. Because of the narrowness of the female pelvic cavity and the size of their heads, human infants are born at an earlier stage in their development compared to most animals and are utterly helpless at birth. So, even after childbirth, human females have to spend months if not years suckling and caring for their offspring.
Although animal protein and fats were a vital component of their diet, the better part of it came from plants— leaves, fruits, roots, tubers etc. These resources are stationary and can be gathered and processed without too much moving around and suited the more sedentary life of women. Children, when they were old enough, along with the elderly, helped with these tasks with the latter passing on the benefit of their experience an some of the skills necessary for survival. Boys likely spent a lot of time hunting down small game, like reptiles, birds and small mammals until they were old enough to accompany their fathers on the hunt. Human societies are characterized the presence of a relatively large contingent of post-menopausal women and elderly men too slow for the hunt (grandparents) who can also lend a hand, sharing some of the workload and helping raise the children. In addition, the wealth of information that they carried in their heads would have been invaluable to the community.

On the hunt in the Kalahari
Hunting was a much more complex and organized activity than simple scavenging and gave the hunter a much better chance of taking away most of the victim’s carcass. Strategy and tactics were an absolute_Fundort_Nariokotome,_Kenia,_Rekonstruktion_im_Neanderthal_Museum.jpg) necessity. Some sort of leadership and assignment of tasks had to be assumed and added brain power would have certainly helped. The skills involved in tracking and understanding animal behaviour took time to acquire and involved a lengthy apprenticeship for the older boys. These same skills could be used to protect the group from the large and dangerous predators that threatened it, especially when they were on the move.
necessity. Some sort of leadership and assignment of tasks had to be assumed and added brain power would have certainly helped. The skills involved in tracking and understanding animal behaviour took time to acquire and involved a lengthy apprenticeship for the older boys. These same skills could be used to protect the group from the large and dangerous predators that threatened it, especially when they were on the move.
Children of a certain age helped with these tasks, acquiring the necessary skills for survival. Boys likely spent a lot of time hunting down small game, like reptiles, birds and small mammals until they were old enough to accompany their fathers on the hunt. Human societies are characterized the presence of a relatively large contingent of post-menopausal women and elderly men too slow for the hunt (grandparents) who can also lend a hand, sharing some of the workload and helping raise the children. In addition, the wealth of information that they carried in their heads would have been invaluable to the community.
Social Organization & Language
These various activities had to be organized and coordinated to a certain extent, which supposes a more sophisticated means of communication than gestures accompanied by grunts, shrieks and screams of apes. There is no physical evidence so we can only surmise, but it is difficult to imagine learning the intricacies of flint working without more detailed instruction. Long practice and much supervision was also required. While apes might learn how to use tools through individual experiment and observation, humans tend to rely more on learning and practice. This becomes more obvious in the production of Mode 2 artefacts. It is possible that the necessary instruction was communicated strictly through ‘signing’ using the hands, however awkward that may have been.

An Aboriginal encampment, near the Adelaide foothills. Alexander Schramm. 1854
Given the diversity of activities undertaken by members of the group, activities that most often took place at a number of different locations, some sort of gathering place was needed where food could be processed and shared out. These acts of sharing (reciprocity) created bonds of trust and a sense of community. The location of the meeting place should be somewhere secure, ideally with trees to provide refuge if needed and near a source of drinking water. Particularly suitable sites would have been used repeatedly over the years, perhaps for days or weeks at a time. These should be identifiable archaeologically by fairly dense concentrations of stone tools and bone but very few have been located.
The development of spoken language provided a unique mechanism for harnessing all the experiences of the group—both collective and individual—and applying them to the solution of long and short term problems. It is certainly true that H. erectus brains were large enough and developed enough to permit speech. Endocasts show significant development of the left hemisphere of the brain, the site of Broca’s Area and Wernicke’s Area, both of which are directly associated with speech and understanding speech.
Homo habilis and H. ergaster managed to coexist for the better part of the Early Pleistocene—probably because they exploited different ecological niches—but, some time around 1.4 million BP, H. habilis disappears from the fossil record followed by the members of the genus Paranthropus at about 1.1 million BP. Mode 1 and Mode 2 stone industries also operate side by side throughout this period and beyond—most likely because they represented different sets of tasks rather than different levels of humanity.
The Middle Pleistocene (ca. 770-126,000 BP)
The Middle Pleistocene sees a profound shift in global climate as the glacial cycles increase from ca. 41 to about 100 thousand years. Within these longer cycles there were frequent, sudden and much more extreme oscillations between cool/dry and warm/moist climates. The glacial phases were much longer, accounting for 90% of a given cycle, while the interglacials were much shorter, lasting about 10,000 years on average.
In Africa, the situation is complicated by the monsoons, which operate on their own 21,000 year cycle. Environmental instability, especially during periods of transition from cold/dry to warm/moist, would have been an enormous stimulus for change in ecosystems all over the continent. These changes were often quite sudden and would have been very challenging for our ancestors. Unfortunately, the fossil and archaeological evidence is very thin and widely scattered for much of this period but it would appear that, by about 600 thousand BP, significant physiological and cultural changes can be detected among early humans. These changes suggest enhanced communication and decision-making skills— it is possible that speech and language developed during this period.
The development of spoken language provided a unique mechanism for harnessing all the experiences of the group—both collective and individual—and applying them to the solution of long and short term problems. It is certainly true that H. erectus brains were large enough and developed enough to permit speech. Endocasts show significant development of the left hemisphere of the brain, the site of Broca’s Area and Wernicke’s Area, both of which are thought by some to be associated with speech and comprehension.
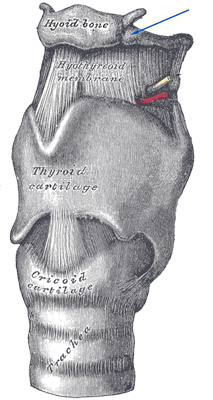
The voice box (larynx) is largely made up of soft tissue which does not fossilize. The only bone is the hyoid which is located at the top of the larynx but, _-_Fig._238.Hyoid-Bone.png) unfortunately, it is not directly attached but is held in place by muscles and so is almost always found on its own. It serves to anchor the tongue and the muscles in the floor of the mouth. Its position in the voice box (high or low) determines the range of sounds produced. The lower the better as far as vocal range is concerned. One fossilized hyoid located high was recovered from a juvenile A. afarensis (c.3.3 million BP) and another, belonging to a Neanderthal from Kebara Cave in the Levant (ca. 60 thousand BP) was located low but nothing has been found in between. Nevertheless, it seems clear that the “power of speech” developed sometime during the Middle Pleistocene but unfortunately, when it comes to fossils, this is a particularly blank period in African prehistory.
unfortunately, it is not directly attached but is held in place by muscles and so is almost always found on its own. It serves to anchor the tongue and the muscles in the floor of the mouth. Its position in the voice box (high or low) determines the range of sounds produced. The lower the better as far as vocal range is concerned. One fossilized hyoid located high was recovered from a juvenile A. afarensis (c.3.3 million BP) and another, belonging to a Neanderthal from Kebara Cave in the Levant (ca. 60 thousand BP) was located low but nothing has been found in between. Nevertheless, it seems clear that the “power of speech” developed sometime during the Middle Pleistocene but unfortunately, when it comes to fossils, this is a particularly blank period in African prehistory.
During the cold/dry phases, there were a number of strategies that a group of hominins might adopt. These included expanding the group’s territory, adapting their food preferences, adjusting the size of the group by splitting it into smaller units in order to exploit more diverse ecological niches, and through technological innovation. Unfortunately, the few examples of hominin fossils that have been found range widely in date as well as in the characteristics they display. Generally speaking, there is a trend among H. erectus/ergaster fossils toward the acquisition of more modern features over time. Their bodies became larger and the proportions were more like our own. More importantly, there was a significant increase in brain size to the extent that there was an overlap with modern humans at the upper end of their range
Homo heidelbergensis/rhodesiensis: The culmination of this evolutionary process saw the emergence of what appeared to be a new species, which was named Homo heidelbergensis—so-called because the first fossil, a mandible dated to about 640 thousand BP, was discovered just outside  that German university town in 1907. Because it had a number of modern features in addition to more primitive ones, it was classified as a new species. Among the primitive features were prominent, arched brow ridges, a wide face and sloping forehead similar to H. ergaster and H. habilis. The derived features, those found among more modern humans, include a significantly larger brain, smaller teeth and a flatter face. Post-cranial finds have been rare but a tibia found at Kabwe suggests that H. heidelbergensis was tall and sturdily built.
that German university town in 1907. Because it had a number of modern features in addition to more primitive ones, it was classified as a new species. Among the primitive features were prominent, arched brow ridges, a wide face and sloping forehead similar to H. ergaster and H. habilis. The derived features, those found among more modern humans, include a significantly larger brain, smaller teeth and a flatter face. Post-cranial finds have been rare but a tibia found at Kabwe suggests that H. heidelbergensis was tall and sturdily built.
At first it was assumed that the species was European, the ancestor of the Neanderthals, but soon a very similar fossil was found at a site known then as Broken Hill (Kabwe) in the then British colony of Northern Rhodesia, now known as Zambia. It was classified as H. rhodesiensis and dated to somewhere between 324,000 and 274,000 BP. Other fossils have subsequently appeared in Africa and Europe—and perhaps in India and China as well. Conventional wisdom has it that, from a single species originating in Africa the line evolved into two branches—a European one that was ancestral to the Neanderthals and an African one that led to modern humans. Today, this seems highly unlikely, especially as another contender , has emerged in Spain, Homo antecessor, as a possible ancestor of Neanderthals at least. Further complicating the issue is the scattered nature (both geographically and temporally) of the few fossils that have been recovered.

Forensic Reconstruction of H. heidelbergensis by Cícero Moraes
At first it was assumed that the species was European, the ancestor of the Neanderthals, but soon a very similar fossil was found at a site known then as Broken Hill (Kabwe) in the then British colony of Northern Rhodesia, now known as Zambia. It was classified as H. rhodesiensis and dated to somewhere between 324,000 and 274,000 BP. Other fossils have subsequently appeared in Africa and Europe—and perhaps in India and China as well. Conventional wisdom has it that, from a single species originating in Africa the line evolved into two branches—a European one that was ancestral to the Neanderthals and an African one that led to modern humans. Today, this seems highly unlikely, especially as another contender has emerged in Spain, Homo  antecessor, which may be an ancestor of Neanderthals at least. Further complicating the issue is the scattered nature (both geographically and temporally) of the few fossils that have been recovered.
antecessor, which may be an ancestor of Neanderthals at least. Further complicating the issue is the scattered nature (both geographically and temporally) of the few fossils that have been recovered.
Coincident with their appearance is the first clear evidence of campsites and the controlled use of fire—both of which indicate a certain level of social organization—along with a marked sophistication in the stone industry. During this period, there are changes to the Acheulean tool-making tradition. The forms remain the same but the finished products are tend to be thinner and more extensively trimmed probably using soft hammers made of antler or bone (left)which offered much more control over the flaking process. The new techniques required more detailed instruction and placed greater demands on the communication skills of the teacher and on the learning of the pupil. A large and more complex brain would have certainly helped.
Homo naledi: The whole issue of the evolution of modern humans became much more complex became much more complicated with the discovery of Homo naledi in 2013. Over 1,500 fossils, belonging to at least 15 individuals and representing both sexes and all age groups were found some 30 metres below ground in the Dinaledi Chamber of the Rising Star cave system about 800 metres from Swartkrans in South Africa. Multiple examples of just about every bone in the body was recovered, many in articulation. They belong to a creature that was slight in build and averaging a little under 1.5 metres (less than 5 feet) tall. Access to the chamber is a tight squeeze for even the slimmest of archaeologists so it is unlikely, to say the least, that they had accidentally fallen in or that the bodies were dragged in by some large predator. In fact, the only other bones found belonged to bird and rodent species rather than game animals. The excavators argue that the remains had been deliberately placed there by other members of the group, possibly as part of some sort of funeral ceremony.
The remains have been dated to 335,000–236,000 years ago, which means they were contemporary with modern humans such as H. heidelbergensis although they are clearly a different species. Like most other members of the genus Homo, they had reduced cheek teeth and reduced post-orbital constriction (a narrowing of the skull behind the eye-sockets) than earlier hominins. Although the size of their brains (ca. 560 cc) was similar to that of australopithecines its configuration was more modern, with a well-developed frontal lobe which has much to do with language, reasoning and social skills. While the arrangement of their shoulder girdle and pelvis are similar to that of australopithecines, their hands and feet were more modern, better suited to manipulating objects and a striding gait.
Homo naledi is only one of a number of “relic” human species that managed to survive along what is considered the main line of human evolution. These include H. floresiensis from the small Indonesian island of Flores and a similarly diminutive hominin from the island of Luzon in the Philippines. They will be dealt with in future articles.
